Research Stories from Ken Cheng's lab
Research stories from Ken Cheng's lab
Bull ants on campus
The research on the night-active bull ants Myrmecia midas is not officially field work because it takes place on the grounds of our campus. But it is field work in that we work in the ants' natural habitat. M. midas is one of a number of species of fierce bull ants in Australia. These ants bite and sting, and scientists working with them take care to handle them carefully.

 The M. midas bull ants come out to forage in the evening twilight. Rush hour is some 20 minutes after sunset. In foraging, they head to a single tree, generally the same one night after night, although not every forager ventures forth every night. The nests are just about all found near the base of a tree, and a minority of foragers, some 30%, simply march up their nest tree. Most stay in their foraging tree for the night, and come home in the wee hours of the morning when the dawn twilight arises.
The M. midas bull ants come out to forage in the evening twilight. Rush hour is some 20 minutes after sunset. In foraging, they head to a single tree, generally the same one night after night, although not every forager ventures forth every night. The nests are just about all found near the base of a tree, and a minority of foragers, some 30%, simply march up their nest tree. Most stay in their foraging tree for the night, and come home in the wee hours of the morning when the dawn twilight arises.
Photos: Left, a night-active bull ant Myrmecia midas at her nest. With large mandibles and eyes and armed with a fierce sting, these ants are ferocious predators. Right, a bull ant Myrmecia midas climbing down a tree. These ants are often found on trees in the twilight because they forage on trees. Photos by Cody Freas.
 In field experiments with M. midas, ants can be captured on their way out foraging or on their way home, and taken for a test under various experimental manipulations, the simplest of which is to displace the test subject to somewhere else in the vicinity. They can also be tested on trees. The ants can be kept content overnight in a well ventilated tube with a bit of food such as honey water, to be tested the next morning, when they are all motivated to get home. Daytime testing has some advantages. For instance, it is easier to spot the test ant, and ants can be filmed with typical video cameras.
In field experiments with M. midas, ants can be captured on their way out foraging or on their way home, and taken for a test under various experimental manipulations, the simplest of which is to displace the test subject to somewhere else in the vicinity. They can also be tested on trees. The ants can be kept content overnight in a well ventilated tube with a bit of food such as honey water, to be tested the next morning, when they are all motivated to get home. Daytime testing has some advantages. For instance, it is easier to spot the test ant, and ants can be filmed with typical video cameras.
Photo: The ants navigate on their nest tree or on their foraging tree. On their nest tree, for example, they generally come around to the side of the tree where their nest is found well before reaching the ground. Orientation is based mostly on the surrounding visual panorama. When the panorama is blocked with black curtains, the ants are disoriented on the tree. This ant in the photo was attempting to place its head in a horizontal position in order to move the head to scan its visual surround. We have observed a number of different ways in which the ants strive to put their heads in an upright position in order to scan. Photo by Cody Freas.
Some references on Myrmecia midas from the lab
Freas, C. A., Narendra, A., & Cheng, K. (2017). Compass cues used by a nocturnal bull ant, Myrmecia midas. Journal of Experimental Biology, 220, 1678–1585.
Freas, C. A., Narendra, A., Lemesle, C., & Cheng, K. (2017). Polarized light use in the nocturnal bull ant, Myrmecia midas. Royal Society Open Science, 4, 170598.
Freas, C. A., Wystrach, A., Narendra, A., & Cheng, K. (2018). The view from the trees: Nocturnal bull ants, Myrmecia midas, use the surrounding panorama while descending from trees. Frontiers in Psychology, 9, 16.
Ferdous, Mst J., Reynolds, A. M., & Cheng, K. (2018). Distinguishing between aapparent and actual randomness: A preliminary examination with Australian ants. Behavioral Ecology and Sociobiology, 72, 113.
Freas, C. A., & Cheng, K. (2019). Panorama similarity and navigational knowledge in the nocturnal bull ant Myrmecia midas. Journal of Experimental Biology, in press.
Desert ant field work
 We work in a semi-natural habitat where red honey ants, Melophorus bagoti, are found, on the outskirts of Alice Springs, Northern Territory. The last 10+ years of field work has been done at a private property south of the town centre, on the way to Alice Springs Airport. The landscape here contains low buildings but also much of the local vegetation, acacia and eucalyptus trees and tussocks of the invasive buffel grass from southern Africa. The distant MacDonell ranges are visible from some spots.
We work in a semi-natural habitat where red honey ants, Melophorus bagoti, are found, on the outskirts of Alice Springs, Northern Territory. The last 10+ years of field work has been done at a private property south of the town centre, on the way to Alice Springs Airport. The landscape here contains low buildings but also much of the local vegetation, acacia and eucalyptus trees and tussocks of the invasive buffel grass from southern Africa. The distant MacDonell ranges are visible from some spots.
Photo: The field site at Alice Springs for studying red honey ants, Melophorus bagoti. The private property has some low buildings, but a lot of natural vegetation as well. Photo by Patrick Schultheiss.
Set ups used in field work
 In much of the experimental ethological work, a feeder stocked with food is set up to lure industrious foragers time and again to come for the goods. We have used mealworm pieces and bits of cookies as rewards. The ants usually like one or the other, or both. Together, they provide the macronutrients of protein, carbohydrates, and lipids. Jokingly, we have called it a balanced diet. We have seen no adverse effects from weeks of such a dietary regime.
In much of the experimental ethological work, a feeder stocked with food is set up to lure industrious foragers time and again to come for the goods. We have used mealworm pieces and bits of cookies as rewards. The ants usually like one or the other, or both. Together, they provide the macronutrients of protein, carbohydrates, and lipids. Jokingly, we have called it a balanced diet. We have seen no adverse effects from weeks of such a dietary regime.
Photo: Feeder used in field research on desert ants, a plastic tub sunk into the ground. A grid is visible too. Photo by Patrick Schultheiss.
The feeder walls are too slippery for the ants to climb out of. Sticks are sometimes placed in the feeder as exit ramps. The exit ramps can be removed to trap ants so that they can be captured for some test. In the photo, an artificial landmark array can also be seen in the distance.
 Some of our artificial landmarks have been more spectacular. We have enclosed a feeder with a round wall carving out an artificial panorama, or set up giant sheets. These artificial panoramas could mimic the natural panorama. Paul Graham, a visiting scientist from the University of Sussex, made an enclosure that mimicked the skyline that the ants saw at the feeder. That is, the top of the black enclosure of sheets and cylinders matched roughly the top of the earthly objects seen at ground level at the feeder, in terms of visual height. The ants used such an impoverished panorama to orient. Or else the artificial surrounds could be new artistic creations with metal or plastic sheets. Multiple studies show that ants learn to orient according to these artificial panoramas.
Some of our artificial landmarks have been more spectacular. We have enclosed a feeder with a round wall carving out an artificial panorama, or set up giant sheets. These artificial panoramas could mimic the natural panorama. Paul Graham, a visiting scientist from the University of Sussex, made an enclosure that mimicked the skyline that the ants saw at the feeder. That is, the top of the black enclosure of sheets and cylinders matched roughly the top of the earthly objects seen at ground level at the feeder, in terms of visual height. The ants used such an impoverished panorama to orient. Or else the artificial surrounds could be new artistic creations with metal or plastic sheets. Multiple studies show that ants learn to orient according to these artificial panoramas.
Photo: In a study led by Antoine Wystrach, a big sheet was used as an experimental landmark near a nest. Photo by Patrick Schultheiss.
 At other times, restrictive channels are used to force the foragers to travel down a particular path. Among other things, the restrictive channel shown in the photo is good for measuring how far the ants run before they turn back. A tape measure on the side of the channel allows easy reading of distances traversed by ants.
At other times, restrictive channels are used to force the foragers to travel down a particular path. Among other things, the restrictive channel shown in the photo is good for measuring how far the ants run before they turn back. A tape measure on the side of the channel allows easy reading of distances traversed by ants.
A less restrictive 'arena' consisting of low (10-cm) walls has also featured in various studies. This arena surrounds a wider area of some 2 m width around the nest and the feeder. It prevents foragers from wandering afield to forage, and induces more foragers to visit our experimental set ups.
Photo: A restrictive channel used to force foragers to travel down a particular path. The rectangular enclosure in the photo usually encloses the nest entrance. At the end of the long channel, the ants typically drop into a feeder provisioned with food. Photo by Patrick Schultheiss.
Wild encounters in field work

 Wild animals can often be seen in the course of field work, such as magpie larks, various Centralian parrots, black kites, and, infrequently, a big wedge-tailed eagle. Lizards could also be spotted occasionally. Even more infrequently, we have spotted snakes, a reason for covering up feet and lower limbs in field work. Snake sightings are rare because all snakes as a rule avoid humans.
Wild animals can often be seen in the course of field work, such as magpie larks, various Centralian parrots, black kites, and, infrequently, a big wedge-tailed eagle. Lizards could also be spotted occasionally. Even more infrequently, we have spotted snakes, a reason for covering up feet and lower limbs in field work. Snake sightings are rare because all snakes as a rule avoid humans.
Photos: Left, a bearded dragon, a common inhabitant of central Australia. Right, a perentie, the largest lizard on the continent.
Some reviews on desert ant navigation from the lab
Cheng, K., Narendra, A., Sommer, S., & Wehner, R. (2009). Traveling in clutter: Navigation in the Central Australian desert ant Melophorus bagoti. Behavioural Processes, 80, 261–268.
Graham, P., & Cheng, K. (2009). Ants use the panoramic skyline as a visual cue during navigation. Current Biology, 19, R935–R937.
Cheng, K., Schultheiss, P., Schwarz, S., Wystrach, A., & Wehner, R. (2014). Beginnings of a synthetic approach to desert ant navigation. Behavioural Processes, 102, 51–61.
Schultheiss, P., Reynolds, A. M., & Cheng, K. (2015). Searching behavior in social Hymenoptera. Learning and Motivation, 50, 59–67.
Wehner, R., Hoinville, T., Cruse, H., & Cheng, K. (2016). Steering intermediate courses: desert ants combine information from various navigational routines. Journal of Comparative Physiology A, 202, 459–472.
Freas, C. A., Fleischmann, P. N., & Cheng, K. (2019). Experimental ethology of learning in desert ants: Becoming expert navigators. Behavioural Processes, 158, 181–191.
Honeybee research
 In the past, Ken Cheng's lab has conducted research on honeybees as well. Much of the work was done on Macquarie University's campus, which boasts large swathes of open green space. Honeybee hives can be kept in a fenced area away from the public. Honeybees can be tested outdoors. In fact, behavioural work is better done outdoors because the environment is more natural and contains more of the cues that the animals use, such as the pattern of polarised light in the sky and the sun itself.
In the past, Ken Cheng's lab has conducted research on honeybees as well. Much of the work was done on Macquarie University's campus, which boasts large swathes of open green space. Honeybee hives can be kept in a fenced area away from the public. Honeybees can be tested outdoors. In fact, behavioural work is better done outdoors because the environment is more natural and contains more of the cues that the animals use, such as the pattern of polarised light in the sky and the sun itself.
Photo: A feeder set up used to attract honeybee foragers. A few worker bees can be seen imbibing the proffered sugar water.
To attract bees, a feeder is set up, offering sugar water. Local bylaws prohibit the use of honey to attract honeybees, but the bees like sugar water. The main attraction is a bottle of sugar water upside down on a plate lined with paper towel. A couple of added features can be seen. A cage outside prevents birds from bothering the bees. The plate sits on a brick immersed in a dish of water, forming a moat. This moat prevents most ants from coming to the sugar feeder, although a few ants have learned to swim the moat, those industrious hymenopterans. (Both ants and bees are hymenopteran insects.)

 Honeybees that are kept for commercial purposes or for research live in constructed hive boxes these days (left). Even amateurs can keep honeybee hives, as the ones in the photograph are on a roof top in suburban Sydney, Australia. Worker honeybees (right) collect nectar and pollen from flowers, and sometimes they collect water. The one in the photo has some orange pollen on her basket—that is the term for the part of the anatomy to which pollen sticks. Many worker bees work from sunrise to sunset, especially when proffered rich sugar water, so that bee scientists could work long, long hours in the summer.
Honeybees that are kept for commercial purposes or for research live in constructed hive boxes these days (left). Even amateurs can keep honeybee hives, as the ones in the photograph are on a roof top in suburban Sydney, Australia. Worker honeybees (right) collect nectar and pollen from flowers, and sometimes they collect water. The one in the photo has some orange pollen on her basket—that is the term for the part of the anatomy to which pollen sticks. Many worker bees work from sunrise to sunset, especially when proffered rich sugar water, so that bee scientists could work long, long hours in the summer.
Photos: Left, honeybee hives on a roof top in a suburb of Sydney, Australia, author: Aussiestarship44. Right, a worker honeybee with pollen on her legs, author: Pollinator at English Wikipedia. Photos from Wikimedia creative commons.
Set ups used in honeybee experimentation
 To get honeybees to work on a task, the experimenter goes to the feeder, and tries to cajole a potential test subject onto a dish of even sweeter sugar water. Once the bee is sucking up the super treat, the experimenter moves her to the test table nearby. After taking her fill, and before zooming home, such a worker typically takes a few good looks at her surrounds, to learn the scenery in order to return later to the great source of food. At the test table, the experimenter could then set up her stimulus array, different odours, coloured cards, or a landmark array. Typically, the rest of the test subject's mates don't know about the super rich set up at the experimental table, and keep going to the feeder. The experimenter can, mostly, train and test her experimental subject unmolested.
To get honeybees to work on a task, the experimenter goes to the feeder, and tries to cajole a potential test subject onto a dish of even sweeter sugar water. Once the bee is sucking up the super treat, the experimenter moves her to the test table nearby. After taking her fill, and before zooming home, such a worker typically takes a few good looks at her surrounds, to learn the scenery in order to return later to the great source of food. At the test table, the experimenter could then set up her stimulus array, different odours, coloured cards, or a landmark array. Typically, the rest of the test subject's mates don't know about the super rich set up at the experimental table, and keep going to the feeder. The experimenter can, mostly, train and test her experimental subject unmolested.
Photo: A honeybee working on an experimental task, trying to choose the correct dish that contains rich sugar water as opposed to unappetising tap water.
 The outdoor lab does not have to be far from one's office building. The tables in this photo show locations for experimenting on honeybees. In fact, in one study (Cheng, 2005), the honeybees learned to do different and conflicting tasks at the two locations located metres apart. Honeybees can learn a lot of different tasks, and they can use an array of contextual cues to differentiate which task they have to do. For instance, they can learn to search on the east side of a cylindrical landmark at one table, but to search at the west side of the same landmark at the other table.
The outdoor lab does not have to be far from one's office building. The tables in this photo show locations for experimenting on honeybees. In fact, in one study (Cheng, 2005), the honeybees learned to do different and conflicting tasks at the two locations located metres apart. Honeybees can learn a lot of different tasks, and they can use an array of contextual cues to differentiate which task they have to do. For instance, they can learn to search on the east side of a cylindrical landmark at one table, but to search at the west side of the same landmark at the other table.
Photo: tables outside an office building that hosted experimental set ups for testing honeybees.
Selected references from the lab on honeybees
Cheng, K. (2000). How honeybees find a place: Lessons from a simple mind. Animal Learning & Behavior, 28, 1–15.
Cheng, K. (2000). Shepard’s universal law supported by honeybees in spatial generalization. Psychological Science, 11, 403–408.
Cheng, K. (2005). Context cues eliminate retroactive interference effects in honeybees (Apis mellifera). Journal of Experimental Biology, 208, 1019–1024.
Cheng, K., & Wignall, A.E. (2006). Honeybees (Apis mellifera) holding on to memories: Response competition causes retroactive interference effects. Animal Cognition, 9, 141–150.
Wignall, A.E., Heiling, A.M., Cheng, K., & Herberstein, M.E. (2006). Flower symmetry preferences in honeybees and their crab spider predators. Ethology, 112, 510–518.
Prabhu, C., & Cheng, K. (2008). One day is all it takes: circadian modulation of the retrieval of colour memories in honeybees. Behavioral Ecology and Sociobiology, 63, 11–22.
Andrew, S. C., Perry, C. J., Barron, A. B., Berthon, K., Peralta, V., & Cheng, K. (2014). Peak shift in honey bee olfactory learning. Animal Cognition, 17, 1177–1186.
Ushitani, T., Perry, C. J., Cheng, K., & Barron, A. B. (2016). Accelerated behavioural development changes fine-scale search behaviour and spatial memory in honey bees (Apis mellifera L.). Journal of Experimental Biology, 219, 412–418.
Developing self control in university students
The lab took a different turn when a talented psychology student and former hairdresser, Megan Oaten, currently at Griffith University, recruited first-year university students to partake in programs of self control. Self control or self regulation means taking actions to achieve long-term goals, often while suppressing the potent impulse of immediate temptations. Oaten had students sign up for weeks or months of a regular exercise regime, a planned study program, or a financial monitoring routine to keep track of and control spending. Students participating in these studies were supposed to report to Megan Oaten regularly, such as every two weeks.
 All these programs led to successful accomplishments by first-year students. Not only did the students stick to their programs, that is, comply with the experimental manipulations, but they also reported improvements in a host of other enterprises of self control, from not losing their temper to eating more healthy food to even less often leaving dishes unwashed in the sink. The most remarkable statistic was the dropout rate once these students had started: 0%! That is unheard of in first-year students. To our knowledge, while the papers have been much cited, this work has not been replicated at all. Given the remarkable success reported in the studies, replications are very much called for.
All these programs led to successful accomplishments by first-year students. Not only did the students stick to their programs, that is, comply with the experimental manipulations, but they also reported improvements in a host of other enterprises of self control, from not losing their temper to eating more healthy food to even less often leaving dishes unwashed in the sink. The most remarkable statistic was the dropout rate once these students had started: 0%! That is unheard of in first-year students. To our knowledge, while the papers have been much cited, this work has not been replicated at all. Given the remarkable success reported in the studies, replications are very much called for.
Photo from Pixabay.
References on self control
Oaten, M., & Cheng, K. (2005). Academic stress impairs self-control. Journal of Social and Clinical Psychology, 24, 254–279.
Oaten, M., & Cheng, K. (2006). Improved self-control: The benefits of a regular program of academic study. Basic and Applied Social Psychology, 28, 1–16.
Oaten, M., & Cheng, K. (2006). Longitudinal gains in self-regulation from regular physical exercise. British Journal of Health Psychology, 11, 717–733.
Oaten, M., & Cheng, K. (2007). Improvements in self-control from financial monitoring. Journal of Economic Psychology, 28, 487–501.
Small-scale spatial cognition in pigeons
Homing pigeons are known for homing from long distances. But a long time ago in Canada, Ken Cheng took on a program of research on how lab pigeons, white domesticated pigeons, find a spot in an arena, a much smaller scale of navigation.
 Pigeons can learn to search for buried food in an arena such as this, in the middle of a typical lab room with benches and doors and tables. Various landmarks can be set up as localisation cues, such as this blue stripe on one of the walls. The food would be in a dish at a constant location in the arena. In this particular arena, the food 'dish' was an indentation drilled into the floor of the wooden arena. The arena is filled with sawdust, which covers the food and sets up a searching task.
Pigeons can learn to search for buried food in an arena such as this, in the middle of a typical lab room with benches and doors and tables. Various landmarks can be set up as localisation cues, such as this blue stripe on one of the walls. The food would be in a dish at a constant location in the arena. In this particular arena, the food 'dish' was an indentation drilled into the floor of the wooden arena. The arena is filled with sawdust, which covers the food and sets up a searching task.
Photo: An arena used for studying small-scale spatial search in pigeons (Cheng, 1988). The food that the pigeons were searching for was buried under the sawdust, in this particular study, 20 cm in front of the blue stripe.
To train pigeons, one exposes the food in its dish at first. Over repeated trials, the dish can be gradually covered more and more, until the sawdust completely covers the few grains, and looks smooth and non-distinctive.

 These low-quality black-and-white images are taken from the videotaped record of pigeons, in the days when videos were recorded on the now extinct videocassettes. Frame-by-frame analyses were done on a monitor with a videorecorder playing the videocassettes. Such were the dark days of the 1980s.
These low-quality black-and-white images are taken from the videotaped record of pigeons, in the days when videos were recorded on the now extinct videocassettes. Frame-by-frame analyses were done on a monitor with a videorecorder playing the videocassettes. Such were the dark days of the 1980s.
Photos: Left, stills from video records of a pigeon searching. On the right side, the blue stripe was in its usual training position, while on the left still, the blue stripe had been moved some distance to the left (looking at the wall on which the stripe was hung). The search of the pigeon shifted to the left, but not by the full extent of the shift of the landmark (Cheng, 1988). Right: a close-up shot of a pigeon working away at the search task. The top of its head was marked with a non-toxic black stripe for ready identification of the front of the head on the video image for data crunching.
Model of small-scale landmark use in pigeons
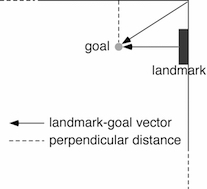 Much of the program of research was done in collaboration with comparative psychologist Marcia Spetch, who won the career research award in 2017 from the Comparative Cognition Society. After more than a decade of research, Cheng, Spetch, and colleagues summarised the state of the art that the pigeon spatial-cognitive research arrived at (Cheng, Spetch, Kelly, & Bingman, 2006). In their model, the pigeon encodes two kinds of metric information: 1) vectors from points of space, such as corners or landmarks that can be conceived as point sources, 2) perpendicular distances from long surfaces. The perpendicular distance representation contains, like vectors, a distance component, but unlike vectors, has no defined starting point. The vectors, on the other hand, point from one particular location to a goal at a particular distance and direction from the starting point, much like classic algebraic vectors. A weighted average of all encoded vectors and perpendicular distances serves well to explain the body of data.
Much of the program of research was done in collaboration with comparative psychologist Marcia Spetch, who won the career research award in 2017 from the Comparative Cognition Society. After more than a decade of research, Cheng, Spetch, and colleagues summarised the state of the art that the pigeon spatial-cognitive research arrived at (Cheng, Spetch, Kelly, & Bingman, 2006). In their model, the pigeon encodes two kinds of metric information: 1) vectors from points of space, such as corners or landmarks that can be conceived as point sources, 2) perpendicular distances from long surfaces. The perpendicular distance representation contains, like vectors, a distance component, but unlike vectors, has no defined starting point. The vectors, on the other hand, point from one particular location to a goal at a particular distance and direction from the starting point, much like classic algebraic vectors. A weighted average of all encoded vectors and perpendicular distances serves well to explain the body of data.
Photo: A schematic drawing of what a pigeon encodes to locate an unmarked spot (labelled goal) in an arena. The bird encodes vectors from distinctive positions in space (a corner, the middle of a stripe, etc.) and perpendicular distances from surfaces. Drawing by Ken Cheng.
Some key references
Cheng, K. (1988). Some psychophysics of the pigeon’s use of landmarks. Journal of Comparative Physiology A, 162, 815–826.
Cheng, K. (1989). The vector sum model of pigeon landmark use. Journal of Experimental Psychology: Animal Behavior Processes, 15, 366–375.
Spetch, M.L., Cheng, K., & Mondloch, M.V. (1992). Landmark use by pigeons in a touch-screen spatial search task. Animal Learning & Behavior, 20, 281–292.
Spetch, M.L., Cheng, K., & MacDonald, S.E. (1996). Learning the configuration of a landmark array I: Touch‑screen studies with pigeons and humans. Journal of Comparative Psychology, 110, 55–68.
Spetch, M.L., Cheng, K., MacDonald, S.E., Linkenhoker, B.A., Kelly, D.M., & Doerkson, S.R. (1997). Learning the configuration of a landmark array in humans and pigeons II: Generality across search tasks. Journal of Comparative Psychology, 111, 14–24.
Cheng, K., Spetch, M.L., Kelly, D.M., & Bingman, V.P. (2006). Small-scale spatial cognition in pigeons. Behavioural Processes, 72, 115–127.
Geometry and navigation
 An even longer time ago, in a paper published in 1986, Ken Cheng reported on a curious error made by rats in relocating a desired location in a rectangular box decked out with various decorations on the walls, and including distinct smells of anise and peppermint in two of the diagonally opposite corners. The much cited paper has led to a plethora of research on the topic, in a range of animals including humans, monkeys, birds, toads, and bees and ants.
An even longer time ago, in a paper published in 1986, Ken Cheng reported on a curious error made by rats in relocating a desired location in a rectangular box decked out with various decorations on the walls, and including distinct smells of anise and peppermint in two of the diagonally opposite corners. The much cited paper has led to a plethora of research on the topic, in a range of animals including humans, monkeys, birds, toads, and bees and ants.
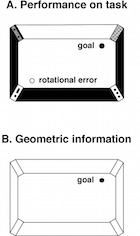
Photo: An illustration of the rotational error made by rats, and young children under some circumstances. The task (A) is to return to an unmarked spot (filled circle labelled goal), using the cues in the rectangular arena after the subject has been disoriented. It is often called a reorientation task. The rats frequently search correctly at the goal location, but they also search at high levels at the spot marked as "rotational error". This is located at 180 degrees rotation from the goal location through the centre of the space. To make that error systematically, the animals must have used only something like the information shown in B, a geometric outline of the space. Drawing by Ken Cheng
When rats, or sometimes 18-24 month old children for that matter, have the task of relocating an unmarked goal in a rectangular space, they search as often at the correct location as at the geometrically equivalent location marked rotational error (A). The rotational error is located at 180 degrees rotation through the centre from the correct location. To do this, rats and young children must have used only the information contained in the shape of the space (geometric information) some of the time (B). What this information consists of is a continuing matter of debate.
 Photo: Photographs of the arena that was used in the original 'geometry' paper (Cheng, 1986), taken outdoors and not under laboratory conditions. In the lab, the arenas would be lit from above by a dim light at the centre of the space, and be filled with sawdust on its bottom, used for hiding rectangular dishes of food. Note the distinctive panels in the four corners, each with a different number of small lights shining through small holes in the panels, with anise and peppermint emanating from two of the corners.
Photo: Photographs of the arena that was used in the original 'geometry' paper (Cheng, 1986), taken outdoors and not under laboratory conditions. In the lab, the arenas would be lit from above by a dim light at the centre of the space, and be filled with sawdust on its bottom, used for hiding rectangular dishes of food. Note the distinctive panels in the four corners, each with a different number of small lights shining through small holes in the panels, with anise and peppermint emanating from two of the corners.
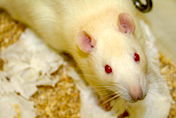 The rats used in the Cheng (1986) study were male Sprague Dawley rats, which are white rats. Males were chosen because they were less variable from day to day. Females are sometimes avoided in rat research because they rotate through an estrus cycle. This procedural detail turned out to matter because subsequent research has shown that male and female rats behave differently in such tasks. This gender bias in animal research has now been seriously criticised. Male rodents vary greatly in hormone levels just like female rats, for example, testosterone level between dominant and subordinate males.
The rats used in the Cheng (1986) study were male Sprague Dawley rats, which are white rats. Males were chosen because they were less variable from day to day. Females are sometimes avoided in rat research because they rotate through an estrus cycle. This procedural detail turned out to matter because subsequent research has shown that male and female rats behave differently in such tasks. This gender bias in animal research has now been seriously criticised. Male rodents vary greatly in hormone levels just like female rats, for example, testosterone level between dominant and subordinate males.
Photo: A white rat of the Sprague-Dawley strain. The eyesight of this strain is poor, among rat strains. But it still uses visual cues in navigation. Actually, various modelling exercises have shown that high-resolution eyesight is not needed for navigation. Using a broad panorama of the visual surround, low-resolution vision such as that possessed by most insects will do just fine, and sometimes works better than higher-resolution visual systems. Photo from Wikimedia creative commons, author: Jean-Etienne Minh-Duy Poirrier.
Some key references on the geometry literature published by Ken Cheng
Cheng, K. (1986). A purely geometric module in the rat’s spatial representation. Cognition, 23, 149-178.
Cheng K. (2005). Reflections on geometry and navigation. Connection Science, 17, 5-21 (special issue on navigation edited by K.J. Jeffery).
Cheng, K., & Newcombe, N.S. (2005). Is there a geometric module for spatial orientation? Squaring theory and evidence. Psychonomic Bulletin & Review, 12, 1-23. (A key review paper of earlier work in the literature that is still cited these days)
Cheng, K. (2008). Whither geometry? Troubles of the geometric module. TRENDS in Cognitive Sciences, 12, 355-361.
Cheng, K., Huttenlocher, J., & Newcombe, N. S. (2013). 25 years of research on the use of geometry in spatial reorientation: A current theoretical perspective. Psychonomic Bulletin & Review, 20, 10333-1054. (A review of various theoretical stances on the geometry literature)